问题分诊室
镁氢包衣对大豆萌发和幼苗生长的影响
发布时间: 2024-09-27
利用种子包衣技术将MgH2包衣于大豆,并设置MgO包衣大豆组作阳性对照和无包衣大豆组作阴性对照,本文系统研究了不同处理对大豆发芽势、发芽率、发芽指数,以及幼苗的地上/地下部鲜重、茎长/粗、总根长、侧根数和叶面积等表型性状的影响。结果表明,包衣对大豆萌发和幼苗生长的促进作用从小到大为:无包衣组<MgO包衣组<MgH2包衣组。采用SPSS进行综合分析,结果显示,MgH2-L3包衣大豆组综合得分最高,为最佳包衣配方。MgH2包衣组大豆的最优表现可能归因于镁与氢的协同生物学效应。
种子萌发是植物生命周期最脆弱的阶段,显著影响后续幼苗的生长及农产品的产量和质量。研究结果表明,氢可通过促进营养物质降解供能、提升抗氧化能力、调控植物内源激素水平,显著促进各种农作物的种子发芽和幼苗生长,包括禾谷类、蔬菜、水果和中草药等。种子包衣是为种子发芽和幼苗生长提供保障的一种新型技术,其利用成膜剂/黏合剂将农药、化肥、微量元素等活性成分均匀黏合在种子表面,具有使用方便、省种省药、提高种子田间成苗率、促进幼苗生长等优点。但是,由于氢气极易挥发,如何实现种子包衣氢以提高氢的生物利用率,是研究人员长期关注的重点方向。
氢化镁(magnesium hydride,MgH2)是一种新型镁基固态储氢材料,具有储氢密度高(100g物质中氢质量为7.6)、水解供氢、长效缓释、使用方便等特点,为种子包衣氢提供可靠氢源。研究表明,MgH2具有促进种子萌发和作物生长的作用。陈茹猛等和He等将MgH2直接加入水或霍格兰培养液中,发现MgH2可显著促进水稻/苜蓿种子的萌发、提高抗环境胁迫能力。此外,本课题组在前期研究中发现,将一定量的MgH2均匀拌于栽培土中,可明显促进生菜种子的萌发,提升生菜的营养物质含量。
本研究采用固态储氢材料MgH2,利用包衣技术将MgH2包衣于大豆,并设置镁肥阳性对照组(MgO包衣大豆组)和阴性对照(无处理大豆组)作对比,研究MgH2对大豆萌发和幼苗生长的影响。
1 材料与方法
1.1 材料
试验大豆(Glycine max(L.)Merr.)种子为中黄37。
包衣剂采用的化学试剂为羧甲基纤维素钠(黏度:50~200mPa·s,阿拉丁)、壳聚糖(黏度:100~200mPa·s,阿拉丁)、聚乙烯醇(Mw13000~23000,98%水解,阿拉丁)和黄腐酸(95%,上海吉至生化科技有限公司)。
镁基材料分别采用MgO(99%,唐山威豪镁粉有限公司)和MgH2(98%,上海镁源动力有限公司)。
基质土为泥炭土,德国K牌876配方。蛭石颗粒大小为0.5~1.0mm。
1.2 试验设计
1.2.1 包衣试验设计
1)包衣剂溶液配置。采用4种包衣剂,分别为羧甲基纤维素钠(S)、聚乙烯醇(J)、羧甲基纤维素钠+聚乙烯醇(S-J)和壳聚糖+黄腐酸(K-H),其配置方法如下。(1)羧甲基纤维素钠包衣剂(S):称量1.5g羧甲基纤维素钠,采用100mL容量瓶定容,80℃加热搅拌2h至完全溶解,配置得到15g/L羧甲基纤维素钠溶液,即羧甲基纤维素钠包衣剂(S)。(2)聚乙烯醇包衣剂(J):称量4g聚乙烯醇,采用100mL容量瓶定容,常温搅拌24h至完全溶解,配置得到40g/L聚乙烯醇溶液,即聚乙烯醇包衣剂(J)。(3)羧甲基纤维素钠+聚乙烯醇包衣剂(S-J):将羧甲基纤维素钠包衣剂(S)和聚乙烯醇包衣剂(J)按照体积比为1∶5配置,即得羧甲基纤维素钠+聚乙烯醇包衣剂(S-J)。(4)壳聚糖+黄腐酸包衣剂(K-H):将3g黄腐酸加入50mL水中,在室温下搅拌2h配置为溶液,再加入1.5g壳聚糖,继续搅拌24h,最后采用100mL容量瓶定容,配置含15g/L壳聚糖和30g/L黄腐酸的混合物,即壳聚糖+黄腐酸包衣剂(K-H)。
2)包衣剂种子。挑取籽粒饱满、个头相当的大豆种子100颗,量取包衣剂2mL,利用物理混匀法将包衣剂与大豆种子充分混合,自然晾干后,获得包衣处理的大豆种子。羧甲基纤维素钠包衣、聚乙烯醇包衣、羧甲基纤维素钠+聚乙烯醇包衣、壳聚糖+黄腐酸包衣处理的种子分别标记为S组、J组、S-J组、K-H组。
3)MgH2和MgO包衣种子。分别采用MgH2和MgO与包衣剂混合均匀,对大豆种子进行包衣,具体包衣过程如图1所示。配置优选的包衣剂,在包衣剂中分别加入不同质量的MgH2和MgO,配置3种浓度梯度:Mg质量浓度为3.5、7和14mg/mL的MgH2和MgO包衣剂,分别记为L1、L2和L3。分别量取2mL的MgH2和MgO包衣剂,与100颗种子充分混合,自然晾干后获得不同浓度的MgH2和MgO包衣种子。MgH2组记为MgH2-L1、MgH2-L2、MgH2-L3;MgO组记为MgO-L1、MgO-L2和MgO-L3。未包衣种子作为阴性对照组,记为CK组。
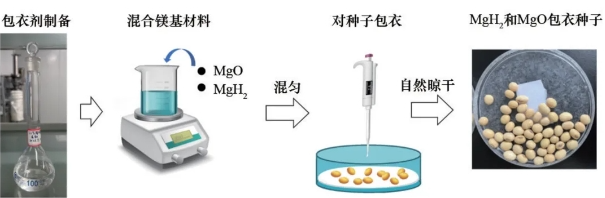
图1 MgH2和MgO包衣大豆种子过程示意
1.2.2 种子萌发试验
利用盆栽法进行种子萌发试验。将不同包衣处理和未包衣种子播于装有基质土与蛭石(体积比为6∶4)的花盆(高15cm,直径10cm)中,每盆播种10颗种子,平行开展3组试验,每盆播种前浇水2L。将花盆置于人工气候培养箱(宁波江南仪器厂,FXA-1000F-4)中,培育条件设为:恒温25℃,12h黑暗/12h光照,光照强度60%,空气相对湿度50%。
1.3 测试方法与数据分析
利用扫描电镜(Scanning electron microscope,ZEISS Sigma 300,Germany)观察MgH2和MgO的表面形貌,并利用扫描电镜照片统计MgH2和MgO的平均粒径;利用光学显微镜(MECATECH 334,France)观察大豆的包衣形态;大豆播种4d开始取样调查大豆的发芽数;利用电子天平(Sartorius SQP,Germany)称量生长7d的大豆幼苗地上部和地下部鲜重;利用根系和叶面积分析系统(TP-PAS-1,China)分析幼苗的茎部/根系形态和叶面积。
发芽率=7d发芽种子数/供试种子数×100%(1)
发芽势=(第4天正常发芽的种子数/处理种子数)×100%(2)
发芽指数=∑(Gt/t)(3)
式中,Gt为在时间t内的发芽种子。
生长7d大豆幼苗的子叶率和真叶率按照公式(4)和(5)进行计算
子叶率=7d幼苗子叶对数/供试种子数×100%(4)
真叶率=7d幼苗真叶对数/供试种子数×100%(5)
利用SPSS27进行单因素方差分析(ANOVA),当P<0.05时差异达显著性水平。利用SPSS27作主成分分析,对单因素变量作特征提取、相关性分析和综合得分。
2 结果与分析
2.1 包衣剂对大豆萌发的影响
为了优选出合适的包衣剂,比较了S组、J组、S-J组和K-H组的发芽率,并对7d大豆幼苗的平均株高、子叶率和真叶率作对比研究,结果见表1。
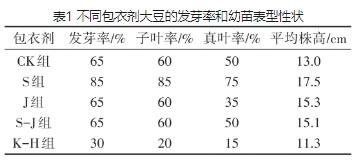
从发芽率来看,S组的发芽率最高,为85%,比CK组的发芽率为65%提高了20%;J和S-J组的发芽率与CK组相同;K-H组的发芽率下降到30%。从子叶率来看,CK组的子叶率为60%,S组的子叶率为85%,比CK组提高了25%;而J和S-J组幼苗的子叶率与CK组持平,K-H组幼苗的子叶率则下降为20%。从真叶率来看,CK组的真叶率为50%,而S组的真叶率为75%,比CK提升25%;S-J组的真叶率与CK组相同;J组和K-H组的真叶率相比CK组反而减低,分别为35%和15%。对平均株高做统计,CK组幼苗的平均株高为13cm,而S组幼苗的平均株高达到17.5cm,相比CK组提高了约35%;J和S-J组幼苗的平均株高为15.3cm和15.1cm,相比CK组分别提高约18%和16%;而KH组幼苗的平均株高降低至11.3cm。
由以上比较分析可知,S组的发芽率和表型性状明显优于CK组,说明羧甲基纤维素钠包衣可促进大豆萌发;而J和S-J组与CK组的发芽率相当,表型性状提升不明显,说明聚乙烯醇和羧甲基纤维素钠+聚乙烯醇包衣剂对大豆发芽影响不大;K-H组的发芽率和表型性状均比CK组差,说明壳聚糖+黄腐酸包衣剂会抑制大豆萌发。因此,后续MgH2和MgO包衣大豆试验优选羧甲基纤维素钠为包衣剂。
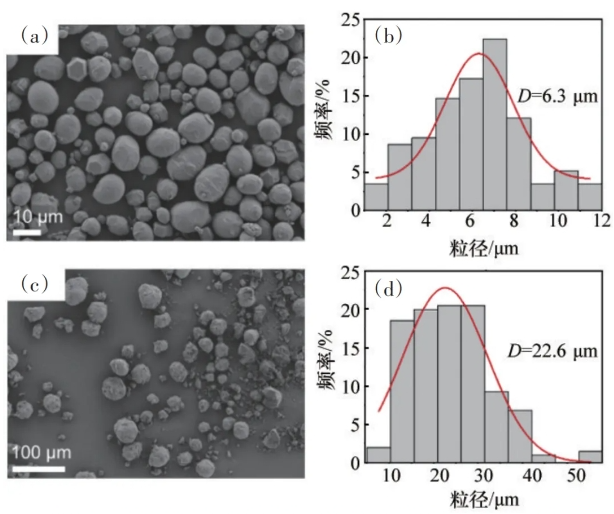
图2 MgH2的SEM图(a)和粒径分析(b),以及MgO的SEM图(c)和粒径分析(d)
进一步采用光学显微镜观察未包衣和不同包衣大豆的表面形貌,结果如图3所示。从图3(a)可以看到,无包衣的大豆表面为纤维网状的种皮结构;而MgO包衣大豆有明显的颗粒附着于种皮上,颗粒数随着MgO浓度的增大而增加(图3(b)~(d));而MgH2包衣大豆则观察到了明显的气泡(图3(e)~(g)),这是由于在包衣过程中,部分MgH2颗粒发生水解反应释放氢气所致。光学显微镜观察结果表明,MgO和MgH2已成功包衣于大豆种皮。
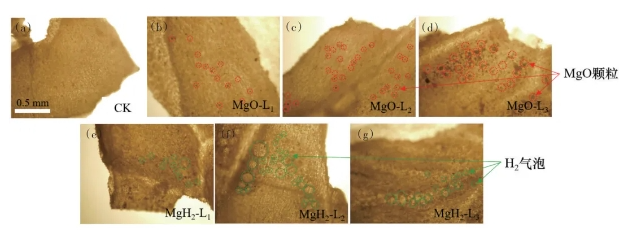
图3 包衣大豆的光学显微镜照片(×10)CK(a)、MgO-L1( b)、MgO-L2( c)、MgO-L3(d)、MgH2-L1(e)、MgH2-L2(f)、MgH2-L3(g)
2.3 镁氢包衣对大豆种子萌发和生长的影响
2.3.1 镁氢包衣对大豆发芽的影响
对不同处理大豆的发芽势、发芽率和发芽指数进行统计分析,结果如图4所示。由图4(a)可知,CK组的平均发芽势为70%,而MgH2组和MgO组的发芽势均有提高,其中MgH2-L3包衣组大豆的发芽势跟CK组相比达到显著差异,平均发芽势达93.3%。发芽率分析结果显示(图4(b)),MgO组和MgH2组的发芽率相比CK均有提高。同时可以发现,MgO包衣组中,只有MgO-L2的发芽率较CK组的发芽率(73%)具有显著差异,而MgH2组的发芽率较CK组均具有显著差异,其中MgH2-L3和MgH2-L1组的发芽率达到100%。发芽指数分析结果显示(图4(c)),MgO组、MgH2组的发芽指数较CK组均有提升,但MgO组与CK组相比差异不显著,而MgH2组与CK组相比差异显著。综上,种子萌发实验结果表明,MgO包衣可促进大豆发芽,而MgH2包衣的促发芽效果达到显著水平。
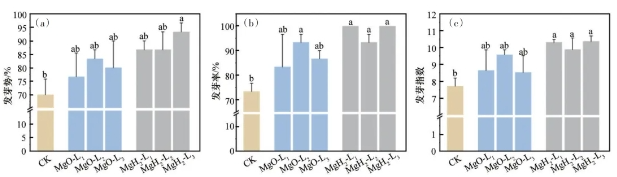
图中数据为平均值和标准误;图中柱子上方的不同小写字母表示相同指标不同包衣材料和质量浓度间差异达5%显著水平
图4 MgH2和MgO包衣对大豆发芽势(a)、发芽率(b)和发芽指数(c)的影响
MgO组和MgH2组的发芽性状较CK组均有提升,这是因为MgO和MgH2包衣处理可为种子提供镁营养,镁具有一定促进大豆萌发的作用。而MgH2组可显著提升大豆的发芽性状,这是因为MgH2包衣处理处在提供镁营养的同时,还可为种子提供氢营养。氢可促进淀粉、蔗糖、脂类等物质分解来为种子萌发提供物质基础,并且氢可调节生长素和赤霉素等内源激素水平来促进种子萌发。MgH2由于镁氢协同作用,促大豆萌发作用较MgO包衣更显著。
2.3.2 镁氢包衣对大豆幼苗鲜重的影响
对不同处理组大豆的地上部、地下部鲜重进行统计分析,结果如图5所示。
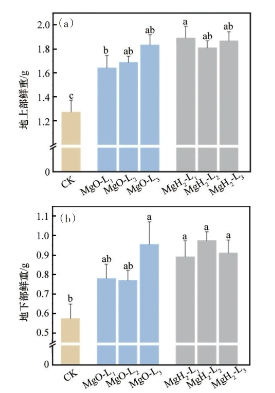
图5 MgH2、MgO包衣对大豆幼苗的地上部(a)和地下部鲜重(b)的影响
从图5(a)中可知,MgH2组和MgO组相较CK组,地上部鲜重均增加并达到显著差异水平。MgO-L1、MgO-L2、MgO-L3大豆幼苗的平均地上部鲜重为1.64、1.69和1.83g,较CK组(1.28g)分别提高了28.1%、32.0%、43.0%。MgH2-L1、MgH2-L2、MgH2-L3大豆幼苗的平均地上部鲜重为1.89、1.81和1.87g,较CK组分别提高47.7%、41.4%和46.0%。
从图5(b)中可知,MgO组和MgH2组相较CK组,地下部鲜重均有增加,但MgO组中只有MgO-L3的地下部鲜重较CK组增加达到显著水平,而MgH2组的地下部鲜重较CK组均显著增加。CK组幼苗的平均地下部鲜重为0.57g,MgO-L1和MgO-L2的平均地下部鲜重为0.78和0.77g,较CK组提高了36.8%和35.1%;而MgO-L3的地下部鲜重较CK组显著提高,达到66.9%。MgH2-L1、MgH2-L2、MgH2-L3的平均地下部鲜重为0.89、0.97和0.91g,与CK组相比分别提高56.1%、70.2%和59.6%。
地上部和地下部鲜重分析结果表明,MgH2和MgO包衣大豆都可显著提升地上部鲜重,但对于地下部鲜重,MgO组仅高浓度MgO-L3增加作用显著,而MgH2包衣大豆3个质量浓度梯度均可显著提升地下部鲜重。这些研究结果提示,镁营养是影响地上部鲜重的主要因素,而镁营养和氢营养可能通过镁氢协同生物效应显著影响地下部鲜重。前期研究结果表明,镁营养元素对叶片中光合效率、叶绿素含量及干物质量均有显著影响,而氢对植物有明显的促生长发育作用,尤其对根系的生长具有明显的促进效应。
2.3.3 镁氢包衣对大豆幼苗生长形态的影响
对不同处理大豆幼苗的7d生长形态进行分析。图6为CK组、MgO组和MgH2组大豆幼苗的平视、俯视和全苗图。从图6可以看出,不同处理组的大豆幼苗生长形态不同。

图6 CK组、MgO组和MgH2组大豆幼苗的全苗、俯视和平视图
对大豆幼苗茎部生长形态(茎长和茎粗)进行分析,结果如图7所示。茎长分析结果显示(图7(a)),MgO组大豆幼苗的茎长较CK组提高,而MgH2组大豆幼苗的茎长较CK组显著提高。CK组的幼苗平均茎长为11.4cm,MgO-L1、MgO-L2、MgO-L3的平均茎长为13.0、12.1和12.6cm,较CK组分别提高14.0%、6.1%和10.5%;而MgH2-L1、MgH2-L2、MgH2-L3的平均茎长分别为13.7、14.2和14.5cm,较CK组分别提高20.2%、24.6%和27.2%。茎粗分析结果显示(图7(b)),MgO组和MgH2组相较CK组的茎粗有提升作用。其中,MgH2-L1的茎粗较CK组显著提升,MgH2-L1的平均茎粗为3.1mm,较CK组(2.7mm)提升了14.8%。因此,从茎部形态来看,MgO包衣对茎长和茎粗的有提升作用但差异不显著,而MgH2包衣主要对提升茎长具有显著效果。因此,MgH2包衣相比MgO包衣对茎部表型性状提升更明显,表明镁氢协同利于大豆茎部发育。
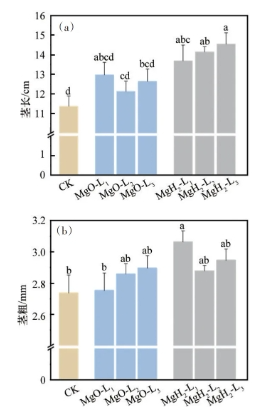
图7 MgH2、MgO包衣对大豆幼苗茎长(a)与茎粗(b)的影响
对大豆幼苗根部生长形态进行分析,结果如图8所示。由图8(a)可知,MgO组和MgH2组均能显著增加大豆幼苗的总根长。CK组的平均总根长为210.4cm,MgO-L1、MgO-L2、MgO-L3的平均总根长为359.9、306.5和397.9cm,较CK组均显著增长。尤其是,MgO-L3的平均总根长相较CK组提升了89.1%。MgH2-L1、MgH2-L2和MgH2-L3的平均总根长为378.3、395.2和389.9cm,相较CK组分别提升了79.8%、87.8%和85.3%。由此可知,MgO和MgH2包衣可显著提升大豆的总根长。由图8(b)可知,MgO组中只有MgO-L3的侧根数较CK组显著增加,而MgH2组的侧根数较CK组均显著增加。CK组的平均侧根数为63.7条,MgO-L1和MgO-L2的平均侧根数为69.0和71.2条,较CK组增加8.3%和11.8%;而MgO-L3的平均侧根数达85.1条,较CK组显著提高到33.6%。而MgH2组的平均侧根数显著增加,MgH2-L1、MgH2-L2、MgH2-L3的平均侧根数为79.0、76.3和78.3条,与CK组相比分别增加24.0%、19.8%和22.9%。图8(c)~(e)为不同处理大豆幼苗的根部照片。可以看到,MgH2组的二级侧根相较CK组和MgO处理组明显发达。二级侧根的增加有利于植物吸收水分和营养物质,从而促进大豆的生长发育。
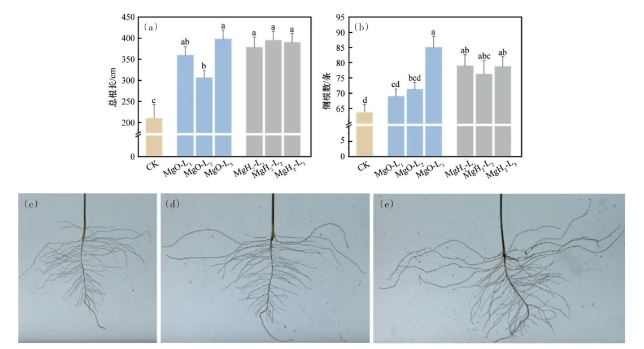
图8 MgH2、MgO包衣对大豆幼苗总根长(a)、侧根数(b)的影响,以及CK(c)、MgO-L2(d)、MgH2-L2(e)大豆根系照片
MgO组和MgH2组的根部发育较CK组提升,这是因为MgO和MgH2包衣处理可为种子提供镁营养,具有促进大豆根系发育的作用。研究表明缺镁会显著降低大豆根系的总根长,而镁处理会显著增强根系的活力和侧根的生长。MgH2包衣对大豆的总根长和侧根数均显著提升,而MgO组对侧根数的提升差异不显著,这是因为MgH2包衣处理为大豆幼苗提供镁营养的同时,还可提供氢营养。氢处理可调控内源H2、H2O2、谷胱甘肽等生长因子促进侧根的发育,这些因素可能导致MgH2包衣显著促进一级和二级侧根的生长。因此,MgH2由于镁氢协同作用,促大豆根系发育作用较MgO包衣更明显。
对大豆幼苗的叶面积进行分析,结果如图9所示。由图9可知,MgO组的叶面积较CK组增加,而MgH2组的叶面积较CK组显著增加。CK组的平均叶面积为553.0mm2,MgO-L1、MgO-L2、MgO-L3的平均叶面积为718.8、678.4和748.7mm2,较CK组均提升。而MgH2组的平均叶面积较CK组显著提升,其中,MgH2-L2的平均叶面积为1169.0mm2,相较于CK组提高了111.4%,而MgH2-L1和MgH2-L3的平均叶面积分别提高60.0%和83.7%。因此,MgH2包衣显著提高了幼苗的叶生长。幼苗的叶生长分析结果表明氢和镁协同促进了叶的生长发育。镁和氢的协同作用有利于调控叶绿素、气孔导度、光化学电子传递速率等来增强叶片的光合作用和蒸腾作用。
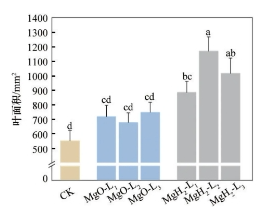
图9 MgH2、MgO包衣对大豆幼苗叶面积的影响
2.4 镁氢包衣大豆的相关性主成分分析
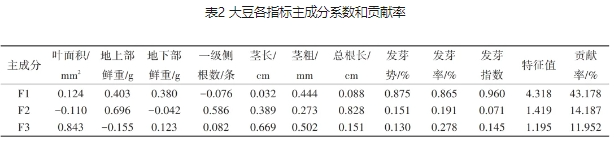
运用SPSS软件对大豆所测得的所有指标进行主成分分析,并计算特征值、特征向量和贡献率,如表2所示。前3个主成分F1—F3的累计方差贡献率达到69.317%,包含了原始变量的大部分信息,也降低了变量个数。3个主成分的特征值分别为4.318、1.419和1.195。F1的特征向量中较大的分量是发芽势、发芽率和发芽指数,说明F1可能代表了影响种子发芽的因素,其贡献率为43.178%,说明发芽因素对包衣大豆种子的萌发性状贡献较大。F2的特征向量中总根长具有高载荷(0.828),说明F2可能代表影响植物根部发展的因素,其贡献率为14.187%。F3特征向量中叶面积具有高载荷(0.843),这表明F3成分与植物的叶面积相关性较高。
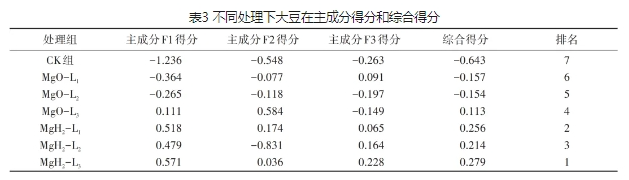
对筛选的主成分进行计算,综合得分见表3。由表3可以看出,MgH2组的综合得分高于MgO组,CK组的综合得分最低。其中,综合得分第1的是MgH2-L3,MgH2-L1和MgH2-L2的综合得分排名分别是2和3。MgO组中,MgO-L3、MgO-L2和Mg-L1的综合得分排名分别为4、5、6。因此,综合得分排名分析表明,MgO包衣可促进大豆萌发和幼苗生长,MgH2包衣比MgO包衣促大豆萌发和生长效果更显著,而MgH2-L3是包衣大豆的最佳配方。
3 讨论
MgH2和MgO包衣大豆促萌发的机理示意图如图10所示。在土壤中,水和微生物的作用下羧甲基纤维素钠包衣剂首先分解/破裂,MgO缓慢水解产生Mg2+,反应式为MgO+H2O→Mg(OH)2,为植株生长提供镁元素;而MgH2缓慢水解产生Mg2+的同时还产生H2,反应式为MgH2+2H2O→Mg(OH)2+2H2。镁对植株生长、养分吸收和产量增加都有明显的促进作用。适宜的镁营养能增加叶片同化物的合成、促进光合产物的运输与分配,进而促进生长和保障产量。同时,氢对植物发芽和幼苗生长具有明显的促进作用。研究表明氢可延缓细胞死亡,原位降低O2⋅-、⋅OH活性氧基团和脂质过氧化,提升细胞活力。同时,氢可调节NO、H2O2等信号分子来调控赤霉素、生长素、细胞周期素等生长因子水平,从而促进种子萌发和幼苗生长。因此,本研究中MgH2包衣组的大豆种子的萌发过程中可缓慢水解提供镁和氢元素(图10(a)),而MgO包衣只能为种子提供镁营养(图10(b)),由于MgH2包衣组可发挥镁氢协同作用,从而显著促进大豆的萌发及幼苗生长。
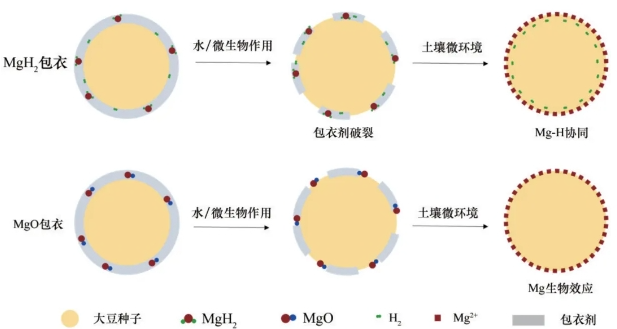
图10 MgH2和MgO包衣大豆促发芽机理示意
4 结论
从发芽率和幼苗表型性状优选出大豆包衣剂,为羧甲基纤维素钠溶液。进一步地,将MgH2包衣于大豆上,成功获得了固态供氢材料包衣大豆种子,显著促进了大豆的萌发,并促进大豆幼苗生长。
研究表明,MgH2包衣大豆可显著提升大豆的发芽率,其中,MgH2-L3和MgH2-L1组包衣大豆的发芽率达到100%。同时,MgH2包衣大豆显著提升了大豆的地上部和地下部鲜重,并可促进茎部、根系和叶片的生长发育,其侧根数、总根长、叶面积较CK组显著提升。而MgO包衣也可提升大豆的萌发和生长,但仅可显著提升地上部鲜重和总根长。最后,对其进行主成分和综合得分分析,结果显示,MgH2-L3包衣大豆组综合得分最高,为最佳包衣配方。由本研究结果可知,MgH2包衣显著促进大豆萌发与幼苗生长,并且优于常规镁肥MgO,这可能归因于镁氢的协同生物学效应。
本文作者:杨海燕,张秋雨,潘琪芳,初少华,杨夕佳,韩京龙,周培,丁文江
作者简介:杨海燕,上海市氢科学重点实验室&上海交通大学氢科学中心,上海交通大学材料科学与工程学院,工程师,研究方向为氢科学、氢生物;丁文江(通信作者),上海市氢科学重点实验室&上海交通大学氢科学中心,上海交通大学材料科学与工程学院,上海交通大学轻合金精密成型国家工程研究中心,教授,中国工程院院士,研究方向为氢科学、轻合金。





 下载app
下载app